
Какие волосы редуцированы у оленей
Опубликовано: 17.09.2024
Archive
Волосяной покров
Шерстный (волосяной) покров — характерная черта облика млек опитающих. Волосы — тоже роговое производное эпидермального слоя кожи. Однако развитие волоса в эмбриогенезе начинается иначе, чем чешуи и пера. Плотный участок эпидермальных клеток приоб ретает цилиндрическую форму и не выпячивается наружу, а на оборот, погружается в мезодермальную часть кожи. Крайние клет ки участвуют в образовании влагалища волоса (фолликула) и саль ных желез, а расположенные по центру превращаются в зачаток самого волоса. Клетки зачатка волоса ороговевают .
Шерстный покров состоит из волос различного типа. Основ ные из них — это пуховые волосы, или подшерсток, остевые воло сы, или ость, чувствующие волосы, или вибриссы.
Основу шерстного покрова у большинства видов млекопитаю щих составляет подшерсток. Между пуховыми сидят жесткие остевые волосы. Именно они создают цветность шерстного покрова. У ведущих подземный образ жизни кротов, слепышеи, цокоров остевые волосы вовсе отсутствуют. У многих копытных, ластоно гих подшерсток имеется у детенышей, а затем он сменяется жес ткими остевыми волосами (например, у северного оленя, котика и др ). Шерстный покров периодически меняется: звери линяют. Белк и, песцы, лисицы, кроты сменяют мех дважды в год. Другие же виды линяют только один раз в год: начав менять шерстный покров весной, они полноценно обрастают новой шерстью к осени (суслики). У человека волосы растут всю жизнь, прирастая в день в среднем на 0,35 мм.
Шерстный покров, как и перья, образование полифункцио нальное. Большинство видов млекопитающих окрашено невзрач но, это скрывает их в местах обитания. В редких случаях окрас и длина шерстного покрова отражает половой диморфизм (у неко торых хищных, приматов). Велико значение шерстного покрова в термоизоляции тела. У северных видов его густота и высота существенно меняются по сезонам.
Особую категорию волос представляют вибриссы: очень длин ные жесткие волосы, выполняющие осязательную функцию. Они сидят чаще на голове (так называемые «усы»), на нижней части шеи, на груди, а у некоторых лазающих древесных форм (напри мер, у белки) и на брюхе. В основании волосяного мешка чувству ющего волоса и в его стенках располагаются мякотные нервные волокна, воспринимающие соприкосновение стержня вибриссы с посторонними предметами. Вибриссы снабжены поперечнополосатой мускулатурой и иннервируются на голове ветвями тройничного нерва.
Видоизменениями волос являются щетина, иглы. Роговым про изводным эпидермиса у китообразных являются пластины китового уса.
Перьевой покров
Тело птицы покрыто перьями. Развитие перьев в онтогенезе птиц указывает на их тесную генетическую связь с чешуями рептилий. Как и зачаток роговой чешуи, зачаток пера возникает в виде бугорка соединительнотканного слоя кожи, пок рытого снаружи эпидермисом. По мере роста бугорок отклоняется назад, а его основание опускается в глубь кожи, образуя влагалище будущего пера и его сосочек, обильно снабжае мый кровью, через который происходит питание растущего пер; Эктодермальная часть зачатка, разрастаясь, дифференцируется на продольное утолщение — будущий стерженъ и два продольных киля этого утолщения, которые позднее распадаются на бородки опахала.
Перья у птиц дифференцированы на контурные, пуховые, соб ственно пух, нитевидные, щетинки. Они располагаются на теле не равномерно, а участками — птерилиями. Такое их распределение не ограничивает свободу движения птицы. Перья регулярно сменяются. Перьевой покров — образование многофункциональное. Его окраска скрывает птицу от взора хищника или, наоборот, демон стративна в половых играх. Структурно и механически (распуша ясъ) он участвует в терморегуляции. И наконец, главное — перь евой покров обеспечивает птице возможность движения в воздухе.
Край крыла образован маховыми перьями: первостепенные махо вые прикрепляются к кисти, второстепенные — к предплечью, третьестепенные — к плечу. Стабильное количество 10—12 первостепенных маховых является систематическим при знаком отряда. Несущую плоскость хвоста образуют рулевые пе рья. Их число тоже регламентировано.
Роговые производные кожных покровов
Роговые производные эпидермиса — приобретения наземных позвоночных животных. Их основное назначение — предохране ние тела от иссушения и защита от повреждений. Эмбрионально роговая чешуя закладывается как выпячивание эпидермиса с ме зодермальным сосочком внутри. Такое сочетание тканей сохраня ется и у взрослых животных.
Утолщения рогового слоя есть на теле жаб в виде бородавок или участков сплошного покрова. На теле рептилий образуется сплошн ой покров из роговых чешуй или щитков.
Поверхность тела гекконов, агам, хамелеонов, гаттерий по крывает примитивная бугорчатая роговая чешуя. На теле настоя щих ящериц и змей чешуя расположена черепицеобразно или ря дами. Голова змей и многих ящериц покрыта упоря доченными рядами роговых щитков. Их расположение учитывает ся при систематической характеристике видов. Видоизме ненные роговые щитки на хвосте гремучей змеи образуют «погремушку». Роговой покров подвержен смене: у змей, оттор гаясь вначале в области челюстей, он сползает с тела полностью, «чулком», у ящерицы отделяется участками. У некоторых репти лий под роговыми чешуями есть окостенения мезодермального слоя кожи.
Тело крокодилов, панцирь черепах и броненосцев покрыто с поверхности роговыми щитка ми. Щитки и чешуйки сохраняются на ногах птиц, на хвостах некоторых млекопитающих (крысы, бобры и др.).
Видоизменениями рогового покрова следует считать роговые чехлы на челюстях — клювы. Они свойственны черепахам, некото рым вымершим рептилиям, однопроходным млекопитающим и, наконец, всем птицам. Многообразие форм клювов птиц отража ет специфику их кормодобывающей де ятельности.
Роговые структуры располагаются на конечностях — когти, ногти, роговые пе репонки и складки на пальцах птиц, по дошвенные подушечки у млекопитающих, кожные гребешки на лапах приматов, копыта на концах пальцев копытных млекопитающих.
К роговым производным эпидермиса относятся также перьевой покров птиц и шерстный (волосяной) покров млекопитающих.
Производные кожных покровов
Производные кориума — чешуя рыб, покровные окостенения скелета, костные пластины рептилий, костные рога копытных. Остановимся на них подробнее.
Тело современных рыб покрыто костной чешуей. В коже хрящевых рыб образуется плакоидная чешуя. Она состоит из остеодентина — вещества, близкого к дентину зубов. Остеодентин — плотное органическое вещество, не имею щее клеточной структуры. Он формируется в кориуме и отличает ся от кости включением в его слои солей извести. Снаружи зубец чешуи покрыт тонким слоем эмали (витродентином), производным клеток эпидермиса.
На челюстях акул плакоидная чешуя укрупняется и выполняет функцию зубов (кожные зубы). Изношенные зубы сбрасываются и замещаются новыми. На поверхности тела некоторых скатов от дельные чешуи разрастаются в шиповатые структуры.
Тела относительно небольшой группы современных костных рыб (панцирников, многоперов) покрывает ганоидная чешуя — производное только мезодермального слоя кожи. Ее нижние слои строятся из настоящей костной ткани (покровное окостенение), а поверхность формируется из остеодентина, свойс твенного плакоидной чешуе. В ганоидной чешуе остеодентин лежит слоями, нарастая одновременно снизу и сверху.
У современных костистых рыб чешуя костная . Чешуйк и формируются из мезодермальных склеробластов. Они лежат в кожных карманах, черепицеобразно налегая друг на друга. Костные чешуйки могут быть округлыми (циклоидныё) или зазубренн ыми (ктеноидныё). Ряды бляшек на теле осетровых рыб и мелкие чешуйки на их теле — это тоже костная чешуя.
Костная и ганоидная чешуя — образования постоянные и не сменяются. На протяжении жизни рыбы чешуи увеличиваются в размерах. Скорость их нарастания неодинакова в разные сезон ы, что послужило разработке методики определения возраста рыбы по шлифам чешуи путем подсчета нарастающих в ней слое в.
Панцирь черепах является уникальным наружным ске летом среди всех наземных позвоночных. Он со стоит из выпуклого овального спинного щита — карапакса и плос кой брюшной пластины — пластрона. По бокам обе части соеди нены костными перемычками. Для головы, конечностей и хвоста панцирь имеет переднее и заднее отверстия.
Наконец, следует упомянуть о рогах копытных — костных вы ростах на лобных костях. Парные костные выросты образуются при участии особого кожного окостенения, которое прирастает к мозговому черепу. Как правило, костные рога отра стают у самцов оленей. К примеру, длина рогов у пятнистого оленя может достигать 93 см. Рога у обоих полов свойственны северному оленю. Костные рога вследствие разрушения костной ткани их основания периодически сбрасываются. Число отростков на рогах измен чиво у разных видов: у пятнистого оленя их в норме 4, может быть 6, марала — 6 — 7.
У полорогих копытных костные выросты на лбу невелики и покрыты постоянными толстыми роговыми чехлами. Уникальным исключением является вилорог , теряющий роговой чехол ежегодно. У жирафа костные рога одеты мягкой кожей. У оленей кожа с волосами покрывает только молодые рога (панты), а затем она отмирает и отпадает.
Строение кожи позвоночных животных
Кожа позвоночных (хордовых) двухслойная: она состоит из наружного эпидермиса и внутреннего кутиса.
Между эпидермисом и следующим, соединительнотканным, слоем кожи — кутисом — лежит базальная мембрана, образованная чаще эпителиальной, но отчасти и соединительной ткан ью.
Эпидермис представлен цилиндрическими клетками и происх одит из эктодермы зародыша. Эпидермис у позвоночных — всегда многослойная ткань, не имеющая кутикулы. У низших позвоночных (круглоротых, рыб, амфибий) эпидермис отграничен от подстилающей его соеди нительной ткани базальной мембраной — бесструктурной переп онкой, которая выделяется сов местно эпидермисом и кутисом . У рыб и личинок амфибий верхний слой эпидермиса (как и у личинок ланцетника) образует на поверхности тонкую пористую каемку, которую считают остатк ом общего мерцательного покрова передковых форм. У современных взрослых амфибий под слоем эпид ермиса обильно ветвятся крове носные капилляры, через них осущ ествляется кожное дыхание (газ ообмен между кровью и окружаю щей средой). Эпидермис богат многоклеточными железами. Их секрет создает на поверхности кожи жидкостную пленку, котор ая препятствует иссушению пок ровов и делает возможным газ ообмен. Благодаря чему амфибии способны выходить на сушу.
Эпидермис рептилий, птиц, млекопитающих — ткань многослойная и достаточно сложная . Его нижний базальный мальпигиев слой в течении всей жизни продуцирует новые слои клеток, поэтому нарушения эпидермиса у них легко восстанавливаются. Верхний слой эпидермиса — роговой — состои т обычно из клеток уплощенной формы. Кровеносных сосудов эпидерм ис не имеет.
Эпидермис дает начало различным роговым производным в виде роговых чешуй и щитков, перьев, волос, копыт, когтей, ногтей, полых рогов, перепонок. В эпидермисе развиваются и разнообразные кожные железы . Наконец, клетки глубоких слоев эпидермиса нередко содержат красящие вещества. Вмест е с пигментами кутиса они обусловливают окраску тела животн ого.
Кутис — внутренний волокнистый слой кожи позвоночных. Его называют также кориум, дерма, собственно кожа. В эмбриоген езе кутис развивается из мезодермального зачатка — кожного листка сомита. Слой кутиса (дермы), как правило, толстый и со ставляет основную часть кожи. В этом слое плотно переплетаются волокна соединительной ткани. Дерма придает коже прочность и эластичность. В кутисе развиваются разнообразные окостенения: чешуя рыб, покровные окостенения скелета, костные рога коп ытных, костные пластины и панцири некоторых рептилий и мле копитающих. У млекопитающих в кутис погружены основания кожных желез, волосяные сумки, у птиц — перьевые фолликулы.
В наружной части кутиса располагаются нервные окончания тепловых и осязательных рецепторов. Здёсь ветвится сложная сеть кровеносных и лимфатических сосудов, находятся пигментные клетки.
Более глубокая часть кутиса имеет рыхлую структуру и служит основным местом образования жировой ткани — подкожной жир овой клетчатки. Здесь накапливается жир — термоизоляционный и питательный материал тела животного. Большой толщины этот слой достигает у морских млекопитающих — китов и дельфинов.
Кожные покровы животных
По кровы животного защищают его тело от механических повреждений, препятству ют проникновению в организм животного бактерий, микроорга низмов, химических веществ. Они ограничивают тело животного и вместе с мускулату рой придают ему форму и удерживают внутренние органы. Покровы участвуют в регуляции вод ного баланса, защищая тело и от потери воды, и от чрезвычайно го ею насыщения. Благодаря наличию специализированных струк тур, желез и обильной гиперемии кровеносными сосудами кожа участвует в дыхании и выведении конечных продуктов.
Кожа и ее производные пигментированы. Окраска тела также многофункциональна: она делает одних животных незаметными, других — демонстративными. Пигменты кожи защищают тело жив отного от избыточного проникновения ультрафиолетовых лучей.
Кожа снабжена нервными окончаниями и специализирован ными рецепторными образованиями, которые осуществляют сенс орную функцию при общении животного со средой. Видимо, нес лучайно большинство органов чувств эмбрионально возникает из специализированных участков наружных покровов зародышей. Производные кожи принимают участие также в формировании органов передвижения (перепонки на лапах, между конечностя ми и телом, маховые и рулевые перья), служат для хватания, борьб ы, защиты (роговые рога, щитки, острые выросты).

- Класс: Mammalia Linnaeus, 1758 = Млекопитающие
- Инфракласс: Eutheria, Placentalia Gill, 1872 = Плацентарные, высшие звери
- Надотряд: Ungulata = Копытные
- Отряд: Artiodactyla Owen, 1848 = Парнокопытные
- Подотряд: Ruminantia Scopoli, 1777 = Жвачные
- Семейство: Cervidae Gray, 1821 = Оленевые, олени, оленьи, плотнорогие
- Род: Alces Gray, 1821 = Лоси
- Род: Capreolus Gray, 1821 = Косули
- Род: Cervus Linnaeus, 1758 = (Настоящие) олени
- Род: Elaphodus Milne-Edwards, 1871 = Хохлатые олени
- Род: Elaphurus Milne-Edwards, 1866 = Олени Давида, китайские олени, милу
- Род: Mazama Rafinesque, 1817 = Мазамы
- Род: Hydropotes Swinhoe, 1870 = Водяные олени
- Род: Moschus Linnaeus, 1758 = Кабарги
- Род: Muntiacus Rafinesque, 1815 = Мунтжаки
- Род: Pudu Gray, 1852 = Пуду
- Род: Rangifer Smith H., 1827 = Северные олени
- Род: Odocoileus Rafinesque, 1832 = Американские олени
- ( в том числе: Род: Ozotoceros Ameghino, 1891 = Пампасные олени
- и Род: Blastocerus Wagner, 1844 = Болотные олени)
Семейство: Cervidae Gray, 1821 = Оленевые, олени, оленьи, плотнорогие

Харктеристика семейства ОЛЕНЕВЫХ. Размеры мелкие, средние, реже крупные. Длина тела 90—310 см, высота в холке 35—233 см. Масса 7—825 кг. Тело стройное с длинными ногами.
У большинства представителей (за исключением Moschus и Hydropotes) самцы имеют рога (у Rangifer рога имеют и самки). В развитом состоянии рога представляют костные, обычно ветвящиеся образования, сидящие на особых выростах (апофизах) лобных костей. Только у оленей родов Elapodus, Mazama и Pudu рога не ветвятся, у всех же остальных они состоят из основного ствола (штанги) и отростков, число которых бывает различно у разных родов и видов оленей и у разных возрастных групп. Самый нижний, отходящий над розеткой отросток рога — первый надглазничный, расположенный над ним — второй надглазничный, третий отросток — средний, а все отростки над ним — верхушечные или венечные. Концы отростков, как и вершина основного ствола, могут вторично ветвиться или уплощаться. Рога один раз в году сбрасываются и отрастают вновь. Растущий рог последовательно проходит стадии соединительнотканную, хрящевую и костную. Снаружи в период роста рога покрыты кожей с короткими волосами: на этой стадии их называют пантами. После окостенения рогов кожа на них отмирает и спадает. В основании рога находится поясок («розетка») с неровной поверхностью, по которому происходит будущая резорбция кости и отпадение рога.
Окраска взрослых особей обычно однотонная, темная, иногда со светлыми пятнами. У примитивных форм и у новорожденных большинства видов оленей окраска пятнистая. У специализированных видов (например, северного оленя) и молодых одноцветная окраска. У многих оленей на задней стороне ляжек расположено светлое пятно — «зеркало». Для волос характерен мощно развитый сердцевинный воздухоносный слой. В кожном покрове имеются разнообразные специфические железы: предглазничные, межпальцевые, плюсневые, хвостовые и др.
Сосков обычно 2 пары. Пальцев четыре, реже два. Боковые пальцы — И и V четырехпалой конечности сильно уменьшены и при ходьбе по твердому грунту не касаются его (исключение представляет северный олень). Боковые пальцы с небольшими копытами.

В черепе пневматизация костей слабая (отмечается лишь у верхнечелюстной, слезной и переднего отдела лобной). На наружной поверхности слезной кости обычно располагается предглазничная ямка специфической кожной железы. В слезной кости два (в виде исключения одно) отверстия слезного канала. Сагиттальный гребень не развит. Этмоидальные отверстия большие. У видов, имеющих рога, лобные кости простираются далеко назад и теменные кости сильно уменьшены. Межчелюстная кость относительно невелика. Костные слуховые барабаны маленькие и соединены с черепом неплотно. Щечные зубы селенодонтные, брахиодонтные. Верхние клыки обычно редуцированы, иногда же развиты хорошо или даже гипертрофированы (у безрогих). Форма нижних клыков сходна с формой резцов. Плюсневые кости боковых пальцев четырехпалой конечности редуцируются, а от пястных сохраняются лишь их верхние или нижние части; иногда исчезают и они. Желудок сложный, четырехкамерный. Желчный пузырь, за редким исключением, отсутствует. Плацента котнледонтная (число котиледонов меньше, чем у Bovidae) или диффузная.
Ареал охватывает Северную, Центральную и Южную Америку (кроме Огненной Земли и Вест-Индии); береговую полосу Гренландии, Северо-Западную Африку; Европу и Азию с прилежащими островами: Шпицберген, Новая Земля, Северная Земля, Новосибирские о-ва; острова Восточной Азии, кроме Курильских и островов Берингова моря; все острова между Азией и Австралией и Шри Ланка.
Населяют в основном леса на равнинах, в предгорьях и горах. Заходят в альпийский пояс и лесостепь. Один вид обитает в тундре. Держатся обычно группами. Питаются различными растениями. Для некоторых оленей характерны миграции. Самки обычно моноэстральные, хотя тропические виды могут быть полиэстральными. Обычно период гона приходится на осень — зиму. Длительность беременности примерно от 5 до 10 месяцев. Число детенышей в помете варьирует от 1 до 4—7.
Американские олени. В Северной Америке, кроме карибу (северного оленя), вапити (местного благородного оленя) и лося, есть еще два вида оленей — олень-мул (запад США, Канады и Северная Мексика) и белохвостый, или виргинский, олень (юг Канады, почти всюду в США, кроме Дальнего Запада, Центральная и север Южной Америки). Кроме того, в Южной Америке 10 (по другим данным — 16) видов оленей, среди них горбатенькие, как кабарга, низкорослые (70 сантиметров в холке) мазамы и пуду-крошки (их рост всего 40 сантиметров). У тех и других рога — простые, неветвящиеся шпильки. В Африке, кроме ланей на северо-западе и благородных оленей, чудом попавших в Сенегамбию, оленей нет. В Азии же около трех десятков видов.
Проделайте теперь мысленное путешествие по ареалу японского пятнистого оленя по имени сика и хуа-лу («олень-цветок»), столь славного высокой ценностью своих пантов. От Уссурийского края вы проследуете по многим восточным провинциям Китая, посетите Корею, Тайвань и Японские острова. Но только мысленный взор покажет вам хуа-лу, а не природа. Потому что повсюду этот олень истреблен. Статистика с посильной точностью утверждает, что за последние сто лет численность пятнистого оленя сократилась «в несколько раз». Сколько сие обозначает, станет понятней, если принять во внимание, что на территории СССР, «даже при наблюдающемся возрастании поголовья к концу пятидесятых годов», можно было насчитать лишь около 1000 этих оленей (лишь половина из них — в прежних местах обитания, прочие — переселенные в заповедники Закавказья, Карельского перешейка и в другие районы СССР). По другую же сторону границы, где возрастания поголовья не наблюдается, голос цифр, вероятно, и совсем унылый. Несколько оживленней выглядит ареал благородного оленя, хотя далеко этому оживлению до толчеи прошлых времен. В Подмосковье на него не поохотишься, потому что он уничтожен там еще до основания Москвы. (В царских угодьях, правда, появлялся и в более поздние времена, но то был завезенный.) В Вологодской, Костромской, Горьковской областях засмеют, если спросите, есть ли в местных лесах олени. А ведь еще в восьмидесятых-девяностых годах прошлого столетия они там водились. Редкостью стал карпатский благородный олень. Мало кавказских и крымских оленей. Мало маралов и бухарских оленей. Немного изюбря на Дальнем Востоке и в Забайкалье. В Китае благородный олень уничтожен полностью, если не считать немногих наиболее находчивых, которые догадались поселиться в священных рощах.
В Америке ареал вапити (близкий вид или подвид благородного оленя, как и марал, изюбрь и прочие названные здесь) какую-нибудь сотню лет назад представлял внушительную площадь, а теперь от него остались три небольших «куска» в Канаде и на западе США, отделенные друг от друга сотнями миль. На Британских островах, Корсике, Сардинии, в Скандинавии всех благородных оленей пересчитать можно. Но и те олени, о которых упоминают сегодня без страха за их будущее, по сути, малые остатки былого великолепия. Говорят, северный олень в Киевской Руси водился. Две тысячи лет назад он упомянут в текстах Цезаря как обитатель Герцинского леса (в Средней Германии и Чехословакии), а еще в прошлом веке прекрасно вписывался в переславльские и новгородские лесные пейзажи. За последние сто лет и в тундрах северных оленей стало меньше в. 15 раз. Упомянутый здесь Ф. Врангель видел стада длиною в 50—100 верст. Где они теперь? Где кавказский лось? Еще в прошлом, жутком для диких зверей веке (таком же, впрочем, как и этот век), когда с недопустимой силой загремели на земле ружейные выстрелы, он жил на Кавказе. Теперь того лося нет. И никто, конечно, не помнит даже, какой масти он был. Когда «как зубья выпадают из гребешка» (тут к месту патетика Маяковского) лучшие из «наших меньших братьев» (весьма точное выражение Есенина), надо бить тревогу!
Волосяной покров – наиболее характерное эпидермическое образование млекопитающих. Отсутствие его у некоторых видов (например, у китообразных) – явление вторичное. Хотя волос – производное эпидермиса, при развитии его зачаток – волосяная луковица – глубоко внедряется в толщу собственно кожи, образуя волосяную сумку. В дно волосяной луковицы вдается сосочек кориума с кровеносными сосудами и нервами. Это так называемый волосяной сосочек. Формирование и рост волоса происходят за счет размножения и видоизменения клеток луковицы, а ствол волоса является уже мертвым образованием, неспособным к росту. В стволе волоса различают три слоя: верхнюю кожицу, корковый слой и сердцевину, Два верхних слоя состоят из плоских ороговевших клеток и, как правило, содержат красящий пигмент. Сердцевина состоит из высохших клеток, наполненных воздухом, что и сообщает шерсти малую теплопроводность. Волосяной покров млекопитающих состоит из волос различного типа. Основными категориями их будут пуховые волосы, ость и чувствующие волосы, или вибриссы. У большинства видов преимущественное развитие получают пуховые волосы, формирующие подшерсток, или пух. Однако у некоторых животных, например оленей, кабанов, многих тюленей, подшерсток редуцирован, и волосяной покров состоит главным образом из остей. Наоборот, у подземных млекопитающих (крота, слепыша, цокора и др.) остевых волос почти нет. У большинства видов волосы на поверхности кожи распределяются неравномерно, собраны в пучки, состоящие из остевого волоса, вокруг которого расположено несколько пуховых (от двух до двухсот). Наблюдается периодическая смена волосяного покрова, или линька. У некоторых видов она бывает дважды в году – осенью и весной; таковы белки, некоторые зайцы, песцы, лисицы. У кротов бывает и третья, летняя линька. Суслики, сурки линяют один раз в году, в весенне-летнее время. У северных млекопитающих по сезонам меняется густота меха, а у некоторых и окраска. Так, у белки на 1 см2 на огузке летом бывает в среднем 4200 волос, а зимой – 8100. На зиму белеют горностаи, ласки, песцы, зайцы-беляки и некоторые другие. Особую категорию волос представляют вибриссы – очень длинные жесткие волосы, выполняющие осязательную функцию. Они расположены на голове, нижней части шеи, груди, а у некоторых лазающих (например, у белки) – и на брюхе. Видоизменениями волос являются щетина и иглы.
Кроме волос, на коже млекопитающих бывают роговые чешуи. Наиболее сильно они развиты у ящеров. Роговые чешуйки наблюдаются также на лапах (мышевидные грызуны) и на хвосте (мышевидные, бобр, выхухоль, некоторые сумчатые). Роговыми образованиями будут полые рога копытных, ногти, когти, копыта. Рога оленей также представляют придатки кожи, но развиваются они из кутиса и состоят из костного вещества.
Мышечная система млекопитающих весьма дифференцирована в связи с разнообразием телодвижений. Есть диафрагма – куполообразная мышца, отделяющая брюшную полость от грудной и имеющая важное значение при вентиляции легких. Хорошо развита подкожная мускулатура. У ежей, ящеров и некоторых броненосцев она обеспечивает свертывание тела в клубок. Эта же мускулатура обусловливает ощетинивание зверей, поднятие игл у ежей и дикобразов, движение вибрисс. На лице – это мимическая мускулатура.
Череп характеризуется относительно крупными размерами мозговой коробки, что естественно связано с большим объемом головного мозга. Кости черепа у млекопитающих срастаются поздно, и это обеспечивает увеличение головного мозга по мере роста животного. Характерно сращение ряда костей в комплексы. Так, четыре затылочные кости образуют одну; сращение ушных костей приводит к образованию единой каменистой кости; сложное происхождение имеют височная и основная кости. Специфической особенностью является строение нижней челюсти, состоящей только из зубной кости. Угловая кость дает начало характерной только для млекопитающих барабанной кости, расположенной в виде яйцевидного вздутия на дне мозговой части черепа. Сочленовная кость, которая у рептилий также входила в состав нижней челюсти, превращается в одну из слуховых косточек среднего уха – молоточек. Нижняя челюсть причленяется к черепу (к височной кости) непосредственно, так как квадратная кость, к которой причленяется нижняя челюсть у рептилий и птиц, также видоизменилась в слуховую косточку – наковальню. В строении позвоночного столба у млекопитающих характерны плоские сочленовные поверхности позвонков и четко выраженная расчлененность позвоночника на отделы: шейный, грудной, поясничный, крестцовый и хвостовой. Первые два шейных позвонка преобразованы в атлант и эпистрофей, а общее число шейных позвонков равно семи. Таким образом, длина шеи у млекопитающих, в отличие от птиц, определяется не числом позвонков, а их длиной. Исключение составляют только ленивцы и ламантины, у которых число шейных позвонков варьирует от шести до десяти. Истинных крестцовых позвонков два, но обычно к ним прирастают еще два хвостовых позвонка. Основой плечевого пояса служит лопатка, к которой прирастает рудиментарный коракоид, и только у однопроходных коракоид представлен самостоятельной костью. Ключица есть у видов, передние конечности которых совершают движения в разных плоскостях, например у обезьян. Виды, перемещающие эти конечности в одной плоскости, например копытные, ключиц не имеют. Конечности млекопитающих вполне типичны для наземных позвоночных, но число пальцев варьирует от пяти до одного. Сокращение числа пальцев или рудиментация крайних из них наблюдается у быстро бегающих видов, например копытных, тушканчиков. Сравнительно медленно перемещающиеся звери, например медведи, обезьяны, опираются при хождении на всю ладонь и ступню (стопоходящие виды); быстрые бегуны, например собаки, копытные, опираются только на пальцы (пальцеходящие виды). Пищеварительный тракт характеризуется большой длиной и хорошо выраженной расчлененностью на отделы. Он начинается преддверием рта, расположенным между мясистыми губами (свойственны только млекопитающим) и челюстями. У некоторых зверей расширение преддверия приводит к образованию больших защечных мешков (у хомяков, бурундуков, сусликов, некоторых обезьян). Мясистых губ нет у однопроходных и китообразных. В ротовую полость открываются протоки слюнных желез, секрет которых не только смачивает пищу, но и химически воздействует (фермент птиалин) на крахмал, превращая его в сахар. Слюна десмодов, питающихся кровью, обладает антикоагулирующим свойством, т.е. препятствует свертыванию крови. У некоторых насекомоядных слюна ядовита и используется для умерщвления добычи. Зубы млекопитающих дифференцированы на группы, в зависимости от типа питания Слабая дифференцировка свойственна малоспециализированным насекомоядным (бурозубкам). У зубатых китов дифференцировка зубов исчезла вторично. Желудок, обладающий многочисленными железами, имеет различный объем и внутреннее строение. Наиболее сложно устроен желудок жвачных копытных, поглощающих огромную массу малокалорийных и трудноперевариваемых кормов. У ящеров и муравьедов зубов нет, а желудок, как и у птиц, состоит из двух отделов: железистого и мускулистого. Сходство усиливается тем, что во втором обнаруживаются умышленно проглоченные камешки, обеспечивающие перетирание пищи. Кроме тонкого, толстого и прямого отделов кишечника, некоторые млекопитающие имеют еще слепой отдел, в котором пища подвергается бактериальному сбраживанию. Особенно сильно слепая кишка развита у кормящихся грубой растительной пищей; длина ее достигает трети длины кишечника. Протоки печени и поджелудочной железы впадают в переднюю часть тонких кишок. Легкие имеют сложное ячеистое строение. Самые мелкие легочные ходы – бронхиолы заканчиваются пузырьками – альвеолами, в стенках которых ветвятся тончайшие кровеносные сосуды. Число альвеол даже у малоподвижных зверей (например, у ленивцев) равно 6 млн., а у весьма подвижных хищных оно достигает 300–500 млн. Механизм дыхания обусловливается изменением объема грудной клетки в результате движения межреберных мышц и диафрагмы. Число дыхательных движений у млекопитающих находится в зависимости от величины животного, определяющей разную интенсивность обмена веществ. Оно составляет (в 1 минуту): у лошади – 8–16, у черного медведя – 15–25, у лисицы –25–40, у крысы – 100–150, у мыши – около 200. Вентиляция легких не только обеспечивает газообмен, но имеет и терморегуляционное значение. При повышении температуры число дыханий увеличивается, а вместе с этим повышается и количество тепла, выводимого из организма. Так, у собаки отношение отдачи тепла при дыхании к общей его потере при температуре воздуха 8° С составляет (в процентах) 14, при 15° С-22, при 30° – 46. Кровеносная система млекопитающих сходна с таковой птиц. Сердце полностью разделено на два предсердия и два желудочка; от левого желудочка отходит одна дуга аорты (но не правая, как у птиц, а левая). Дуга аорты отсылает к голове сонные артерии и, огибая сердце, тянется под позвоночным столбом, по пути направляя разветвленные сосуды к системам органов. Венозная система характеризуется отсутствием воротного кровообращения в почках, которое в печени, как и у других позвоночных, хорошо развито. В печени происходит нейтрализация токсических продуктов белкового обмена. Размеры сердца варьируют в зависимости от величины тела, образа жизни, а в конечном счете – в связи с интенсивностью обмена веществ. Масса сердца, выраженная в процентах к общей массе тела, составляет: у беззубых китов –0,6–1,0, у дикого кролика –3,0, у крота –6,0–7,0, у летучих мышей –9,0–15,0. Все эти особенности обеспечивают млекопитающим более высокий уровень обмена веществ и общей жизнедеятельности.
Прочие статьи:
Обонятельный анализатор
Вид чувствительности, направленный на восприятие различных пахучих веществ с помощью обонятельного анализатора, называется обонянием. Обоняние имеет важное значение в обеспечении безопасности, люди с нарушением обоняния чаще подвергаются .
Географическое распространение и экология обитания
В Черном море обитают 3 подвида азовского пузанка, отличающихся меньшим числом тычинок: С. с. palaeostomi (оз. Палеостоми на Западном Кавказе), С.с. nordmanni (северо-западная часть Черного моря и его реки, например, Дунай), С.с. tanaica .
Этнический и антропологический состав европейской части СНГ
«Этнический состав. Самый многочисленный народ СНГ – русские (130 млн.), широко расселены по всей территории. По происхождению, языку и культуре им родственны украинцы (40,8 млн.) и белорусы (9,2 млн.). Русские, украинцы и белорусы состав .
--> -->Меню сайта -->
--> -->Статистика -->
--> -->Форма входа -->
Эпидермис млекопитающих образует различные роговые производные. В первую очередь, это волосяной покров, который для представителей данного класса столь же характерен, как покров из перьев для птиц или чешуйчатый покров для рептилий. Он выполняет теплоизолирующую функцию, защищает кожу от повреждений, намокания (у полуводных зверей) и некоторых паразитов, улучшает аэро- и гидродинамические свойства тела, обеспечивает видоспецифичность окраски, маскировку и цветовую демонстрацию (для некоторых видов или в определенное время года), служит осязательным рецептором.
У большинства млекопитающих волосы растут на всем теле за исключением губ и участка на конце морды вокруг ноздрей (в этом случае его называют носовым зеркалом), а у некоторых видов — и подошв. Лишь немногие млекопитающие вторично, полностью или частично, утратили мех. В большей или меньшей степени волосяной покров редуцирован у ластоногих, особенно это заметно у моржа (Odobenus rosmarus), в наименьшей мере — у ушастых тюленей (Otariidae), которые более чем другие ластоногие связаны с сушей. Почти полная редукция кожной растительности наблюдается у китообразных; у белухи (Delphinapterus leucas) и нарвала (Monodon monoceros) она совсем отсутствует. Из рукокрылых голой кожей обладает ошейниковая летучая мышь (Chiromeles torquatus), у которой сохранился лишь ошейник из редких волос. Виды, имеющие почти голую кожу, известны и среди грызунов — например, голый землекоп (Heterocephalus glaber).
В период утробного развития у млекопитающих имеется сплошной эмбриональный пух, или лануго (lanugo foetalis), который затем сменяется постоянным волосяным покровом. Важно заметить, что эмбриональным слоем пуха обладают и виды, практически лишенные волосяного покрова во взрослом состоянии (китообразные, слоны, бегемоты), что дополнительно указывает на вторичный характер его утери. 
Схема строения волоса (по Сапину и Биличу, 2007):
1 — волосяная сумка; 2 — кора волоса; 3 — мозговое вещество волоса; 4 — кутикула; 5 — наружное корневое влагалище; 6 — два слоя внутреннего корневого влагалища; 7 — волосяная воронка; 8 — базальный (ростковый) слой эпидермиса; 9 — роговой слой эпидермиса; 10 — сальная железа; 11 — мышца, поднимающая волос; 12 — луковица волоса; 13 — сосочек волоса. В отдельном волосе различают нижнюю вздутую часть — луковицу и выдающийся над поверхностью кожи длинный цилиндрический стержень. Нижняя часть стержня вместе с луковицей образует корень волоса. Нежная ткань луковицы не может удерживать растущий стержень, поэтому внутри нее имеется волосяная сумка с двумя вложенными друг в друга тонкостенными влагалищами. Наружная поверхность сумки образована волосяным мешочком, или фолликулом (folliculus pili) — соединительной тканью, являющейся производным дермы. Затем следует прозрачная стекловидная оболочка. Изнутри волосяную сумку выстилает наружное волосяное влагалище, представляющее собой впячивание росткового слоя эпидермиса. Оно медленно выдвигает из сумки внутреннее влагалище, которое надето на волос и плотно сцеплено с ним благодаря рельефу его внешнего слоя. Ближе к выходу из сумки внутреннее влагалище, состоящее из особого кератина, переваривается симбиотическими бактериями. В дно волосяной сумки вдается очень маленький волосяной сосочек, образованный дермой и служащий местом входа в волос питающих его тончайших капилляров. Ближе к выходу сумка образует свободное щелевидное пространство, куда открывается проток сальной железы (иногда 2-3 желез). Ее секрет которой смазывает волос, сообщая ему большую прочность и водоупорность, а также способствуют сохранению им эластичности. В волосяной сумке расположены и окончания чувствующих нервов, проникающие сюда через волосяной сосочек. К нижней части волосяного мешка снаружи прикрепляются пучки гладких мышечных волокон (musculas arrectores pilorum), сокращением которых волос, имеющий обычно несколько наклонное положение, может быть выпрямлен и приподнят. Это движение и обусловливает ощетинивание, особенно выраженное у животных с длинной шерстью или гривой. 
Структура стержня волоса. В самом волосе различают 3 слоя. Поверхность волоса покрыта тонкой чешуйчатой кожицей, или кутикулой (cuticula), образованной одним или несколькими слоями плоских ороговевших клеток. Они либо опоясывают стержень волоса сплошными кольцами, либо располагаются по поверхности частично перекрывающими друг друга подобно черепице или только соприкасающимися чешуйками. В соответствии с этим чешуйчатый слой называют кольцевидным, черепицеобразным или мостовидным. Чешуйчатый слой защищает волос от разного рода физико-химических воздействий. Под поверхностным слоем расположен плотный корковый слой (cortex) — он состоит из вытянутых по продольной оси волоса и прочно спаянных между собой веретеновидных ороговевших клеток. Этот слой обусловливает крепость и упругость волоса. Центральную часть волоса занимает рыхлая сердцевина (medulla), состоящая из расположенных поперек продольной оси волосяного стержня сплющенных ороговевших клеток, которые разделены воздушными промежутками. В основании и вершине зрелого волоса сердцевина вообще отсутствует и на ее месте образуется канал. Особенно большое количество воздуха содержится в волосах обитателей районов с холодными зимами; это увеличивает теплоизолирующие свойства их волосяного покрова. Морфологическая структура волос у разных групп млекопитающих довольно резко отличается, в частности характерные очертания кутикулярных клеток могут служить четким систематическим признаком. 
Типы строения кутикулярного слоя волоса:
1 — кольцевидный; 2 — черепицеобразный; 3 — мостовидный. Волос млекопитающих — сложное многоклеточное образование, что отличает его от «волоса» беспозвоночных животных, представляющего собой кутикулярный вырост одной клетки. Происхождение волоса остается загадкой, его, в отличие от птичьего пера, не удается вывести из чешуи. Перо проходит в своем развитии стадию выпячивания эпидермиса в виде сравнительно тонкостенного колпачка, заполненного богатой капиллярами мякотью дермы. При закладке волоса клетки эпидермиса перемещаются противоположным образом — они впячиваются в дерму и оставляют на поверхности узкую, но глубокую впадину, на дне которой начинают энергично делиться. По мере своего роста волосяной зачаток все глубже погружается в дерму. Нижняя сторона зачатка вогнута и образует впадину, куда входит развивающийся волосяной сосочек — единственный принадлежащий дерме компонент. Разрастаясь и дифференцируясь, эпидермальный зачаток дает начало основанию волоса — волосяной луковице. Наружные слои зачатка формируют волосяную сумку и сальные железы, из его внутренних слоев образуется собственно волос. Рост волоса происходит за счет интенсивного деления базальных клеток волосяной луковицы. Вершина растущего волоса выдвигается из волосяной сумки. Последовательно оттесняемые кверху, клетки волоса постепенно ороговевают. Они прочно склеиваются между собой, переполняются белком кератином, получают вкрапления пигмента, отмирают и высыхают. Таким образом, весь волос, кроме луковицы, представляет собой мертвое образование. С прекращением роста волоса его луковица зарастает, спадает с сосочка и ороговевает. Наконец, волос выпадает. Новый волос возникает уже на другом сосочке, образующемся, впрочем, в старой волосяной сумке. 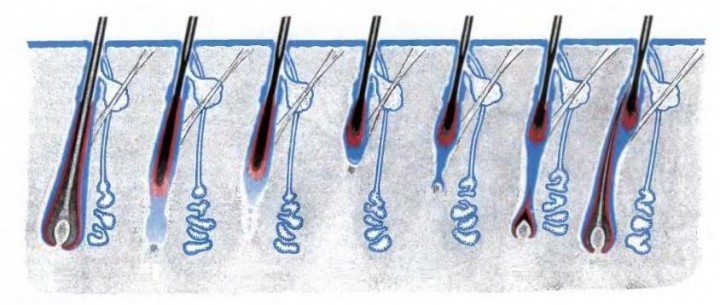
Последовательные фазы роста волоса. Красным цветом отмечено внутреннее эпителиальное влагалище корня, синим цветом — его наружное эпителиальное влагалище. (По Фольмерхаус и Фравейн, 2003.) Волосы млекопитающих неоднородны. В зависимости от гистологического строения, размеров и формы они могут быть разделены на две основные категории: ость и пух. Остевые, или покровные, волосы (шерсть) характеризуются длинным и толстым, прямым или слегка изогнутым стержнем, обычно имеющим в концевой части расширение — гранну. У остевых волос хорошо развита сердцевина, состоящая из нескольких рядов клеток; тип их кожицы черепицеобразный. Помимо остевых волос, у многих млекопитающих имеются реже расположенные, но более длинные направляющие волосы, возвышающиеся над общим меховым покровом. Пуховые волосы (подшерсток) наименьшие по размерам; они очень тонкие, мягкие и многочисленные. Обычно они волнообразно изгибаются и обладают слабо развитой сердцевиной с одним рядом четкообразно расположенных клеток либо совсем лишены ее; кожица у них кольцевидного типа. Волосы подшерстка собраны в более или менее плотные группы вокруг оснований остевых волос. Иногда присутствуют также промежуточные волосы, занимающие переходное положение между остью и пухом. Независимо от принадлежности к той или иной категории волосы весьма разнообразны как по форме стержня, так и по характеру его изогнутости. Волосы, круглые на поперечном срезе, чаще бывают прямыми и торчащими. Волосы, которые овальны или сплющены на срезе, легче сгибаются и нередко образуют волнистый или курчавый шерстный покров. 
Слева — схематическое изображение основных категорий волос:
I — направляющие волосы; II — остевые волосы; III — пуховые волосы; 1 — прикорневая зона; 2 — основание волоса; 3 — переходная зона; 4 — гранна; 5 — кончик волоса.
Справа — пучки волос и двухъярусность шерстного покрова:
1 — пуховые волосы, 2 — остевые волосы. Основные функциональные значения шерсти — предохранение подшерстка и кожи от различных внешних механических воздействий и придание телу определенной окраски. Шерсть защищает животное от палящего солнца и леденящего ветра, способствует постепенному испарению пота в пустыне и предотвращает намокание во время дождя или при нырянии. Вопреки распространенному мнению, водоотталкивающие свойства шерстному покрову (как и перьям птиц) придает не столько гидрофобная жировая смазка, сколько сформированная волосами структура — «решетка». Вдавливаясь в ее ячейки, поверхностная пленка окружающей воды даже при хорошем смачивании ости образует выпуклые по направлению к поверхности кожи мениски (а не вогнутые, как в стеклянных капиллярах), которые поэтому не втягивают, а выталкивают воду. Подшерсток образует эффективный теплоизоляционный слой.
Особенно мощный подшерсток из извитых волос развивается у одетых густым мехом водных млекопитающих — выхухоли (Desmana moschata), бобра (Castor fiber), выдры (Lutra lutra), норки (Mustela lutreola) и др. Возвышающиеся над подшерстком остевые и направляющие волосы имеют копьевидную форму; в воде их верхняя часть отклоняется в противоположную движению сторону и ложится на пружинящий слой пуховых волос. Поэтому волосяной покров этих животных образует пружинящую (демпфирующую) систему, аналогичную упругой коже китообразных. Специальное назначение имеют сильно удлиненные волосы некоторых млекопитающих, образующие хвостовой пучок (служит для обмахивания тела), гриву, баки, бороду (защищающие лицевые части, шею) и т. п. 
Схема положения волос у водных, покрытых мехом млекопитающих на суше (А) и при плавании и нырянии (Б) (по Наумову и Карташеву, 1979):
1 — остевые и направляющие волосы, 2 — пуховые волосы. Для большинства млекопитающих характерна двухъярусность меха, слагающегося из шерсти и подшерстка. При дополнительном наличии промежуточных волос возникает трехъярусность меха. У небольшой части млекопитающих волосяной покров преимущественно или полностью состоит из волос лишь одной категории. Так, кроты (Talpinae) и слепыши (Spalacinae) имеют хорошо развитый подшерсток и почти полностью лишены остевых волос. Наоборот, у взрослых оленей (Cervidae), свиней (Suidae) и настоящих тюленей (Phocidae) подшерсток редуцирован и покров состоит главным образом из остей (у молодых особей этих зверей подшерсток развит хорошо). Отсутствие ярусности (как и безволосость) — результат редукции ранее существовавшего полного волосяного покрова. 
Двухъярусный мех ушастого тюленя отличается большой густотой. Каждый пучок волос, помимо остевого волоска, может включать до 50 пуховых волосков, повышающих теплоизоляцию. (По Уленбрук, 2009.) Волосы обычно располагаются на теле в определенном порядке — либо равномерно поодиночке (например, у коровы (Bos primigenius taurus), лошади (Equus ferus caballus)), либо более или менее сложными группами с различным числом и локализацией отдельных волосков (свиньи (Suidae), хищные и др.). Как правило, волосы закреплены в коже не перпендикулярно ее поверхности, а направлены вершинами в определенную сторону, так что более или менее плотно прилегают к коже. Такой наклон мехового покрова — ворс — способствует обтеканию тела потоками воздуха и воды. Господствующее направление ворса — от головы к хвосту либо от спины к брюху. Характер ворса меняется в тех местах, где кожа часто подвергается образованию складок и растяжению. На некоторых участках шерсть образует завихрения.
Ворс тесно связан с образом жизни и характером движения. Так, у ленивцев, постоянно висящих на деревьях вниз спиной, волосы уложены в направлении от брюха к спине, что можно связать как с силой тяжести, так и с «причесывающим» влиянием дождей. У типичных землероев — кротов (Talpa), слепышей (Spalax), слепушонок (Ellobius) — очень низкий и бархатистый мех расположен вертикально, что дает возможность зверькам быстро двигаться в узких подземных ходах как вперед, так и назад, не оборачиваясь. Сходный волосяной покров имеют обитатели густых травянистых зарослей и подстилки (землеройки). То или иное расположение волос строго специфично для каждого вида млекопитающих и служит хорошим систематическим признаком и показателем филогенетических отношений. 
У двупалого ленивца Гоффмана (Choloepus hoffmanni) волосы уложены в брюшно-спинном направлении. Наибольшего развития волосяной покров достигает у млекопитающих арктических и умеренных областей, для которых его теплоизолирующие свойства имеют особо важное значение. Густой мех имеют некоторые тропические формы (обезьяны, ленивцы), но у них главное значение приобретает защита от проливных дождей и укусов насекомых (безволосые тропические животные — слоны, носороги и др. — хорошо защищены от насекомых грубой толстой кожей). В зависимости от климатических условий, мест обитания и особенностей образа жизни высота волос и степень опушенности отдельных участков тела у разных видов не одинакова. У белок (Sciurus) и других древолазаюших млекопитающих наиболее длинные волосы располагаются на спине, крестце и хвосте, у настоящих норников — сусликов (Spermophilus), сурков (Marmota) — на боках, у лисиц (Vulpes) — на лопатках, у зайцев (Lepus) — на брюхе; у роющих видов длина волос по всему телу одинакова. Имеются различия и в распределении плотности (густоты) волосяного покрова. Снашивающийся мех нуждается в смене, которая происходит либо постепенно, либо в определенное время года, когда в процессе линьки сменяется весь или почти весь шерстный покров. В результате сезонных линек у млекопитающих арктических и умеренных областей зимний мех становится гуще и теплее, а летний оказывается не столь густым. Мех росомахи (Gulo gulo) обладает уникальной особенностью — не покрывается инеем на морозе.
Окраска волос далеко не всегда зависит от окраски кожи. Как правило, в клетках сердцевины и коркового слоя волос содержатся пигменты из группы меланинов. Они поступают из пигментных клеток дермы и в зависимости от комбинации создают различные оттенки коричневого, черного и рыжеватого тонов. Содержащиеся в волосе пузырьки воздуха ослабляют интенсивность окраски; если воздушных полостей много, то темная окраска выглядит серой, а в отсутствие пигмента они создают серебристо-белую окраску. Седение волос при старении обуславливается тем, что клетки сердцевинного слоя частью превращаются в фагоциты, поедают пигмент и, выходят из волоса в кожу, уносят его. Помимо ослабления пигментации, обесцвечивание волос связано с увеличением содержания в них воздуха. (Отметим также, что по истечении определенного времени меланины разрушаются — именно этим обусловливается блеклый окрас дошедших до нас ископаемых волос). Окраска шерсти, как правило, является покровительственной. Она нередко подвержена значительным сезонным изменениям; так, различные арктические животные зимой покрыты белым мехом, хорошо сливающимся со снежным покровом и делающим их незаметными, а перелиняв, покрываются темным летним мехом. Иногда окраска участвует в формировании вторичных половых признаков. Кроме того, окрас подвергается внутривидовой изменчивости. Периодически волосяной покров меняется — происходит линька. 
Различия в рисунке на крупе разных особей зебры Греви (Equus grevyi) — пример внутривидовой изменчивости в окраске. Уход за мехом, от состояния которого зависит, выживет ли зверь, осуществляется с помощью зубов, когтей или ногтей. Так животное избавляется от грязи, колтунов и эктопаразитов, а кроме того, распределяет по волоскам секрет сальных желез. Некоторые млекопитающие производят взаимную чистку не только с целью поддержания гигиены, но и как проявление родительской заботы о потомстве, а также для установления близких контактов в размножающейся паре или иерархии внутри группы.
Видоизменениями толстых и круглых на срезе остевых волос являются щетина и иглы. Щетина — переходный тип между волосами и иглами. Она обычна у млекопитающих, обладающих иглистым панцирем, наибольшего же развития достигает у тенреков (Tenrecidae) и свиней (Suidae). Так, щетина кабана (Sus scrofa) представляет собой прекрасную броню, по которой соскальзывают шипы растений во время передвижения через густые заросли. Иглы образуют своего рода колючий защитный панцирь у ехидн (Tachyglossidae), ежей (Erinaceidae), дикобразов (Histricidae), древесных дикобразов (Erethizontidae) и др.; они характеризуются мощным развитием коркового слоя, врастающего продольными перегородками в сердцевину, прочностью, большой упругостью и очень острой вершиной. Обычно иглы сосредоточены на спинной стороне туловища, тогда как на брюхе имеются грубые волосы. Между ними и настоящими иглами на теле животного можно найти целый ряд переходов. 
Иглистый панцирь обыкновенного ежа (Erinaceus europaeus). вперед
Семейство отр. парнокопытных, относится к жвачным. Разделяется на 3-4 подсемейства, включает 14-16 родов, около 40 видов; на территории России 4 рода и 6-7 видов, относящихся к 2 подсемействам. Иногда сюда включают сем. кабарговых. Известны с нижнего олигоцена. 
Оленьи — красивые животные, чаще всего крупные (лоси весят более полутонны), но есть и такие, как южноамериканские мазамы, размером со среднюю собаку (вес до 25 кг). В целом для семейства показатели таковы: длина тела 90-310 см, масса 7-850 кг. Большинство стройные, высоконогие, с длинной шеей, короткохвостые. Голова удлинённая, на конце морды обычно голый участок — носовое зеркало.

У самцов большинства оленьих имеются рога — чаще всего ветвистые костные выросты лобных костей черепа; 
самки безрогие (за исключением северного оленя). У мелких оленей из тропических лесов (мунтжаки из Юго-Вост. Азии, южноамериканские пуду) рога небольшие, с 1-2 отростками, у оленей из северных краев огромные — до 20-30 отростков. У вымершего большерогого оленя расстояние между вершинами рогов достигало 3 м! У лося рога уплощенные, лопатообразные. Рога ежегодно сбрасываются весной и вырастают вновь осенью ко времени гона; на оставшемся основании рога, как и на черенке опавшего листа, появляется утолщение — "розетка". Во время роста рога мягкие, пронизаны кровеносными сосудами, покрыты кожей с короткими бархатистыми волосами с короткой бархатистой шерстью. Такие растущие рога называют пантами.

Пястные и плюсневые кости конечностей срощены в цевку. Из пальцев хорошо развиты только 3-й и 4-й; боковые пальцы обычно редуцированы, при ходьбе не касаются субстрата.
Волосяной покров густой, с сердцевинным воздухоносным слоем. Имеется большое количество кожных желез — 
межпальцевые, плюсневые, хвостовые, предглазничные и др. Основной фон окраски рыжевато-бурый; у примитивных оленьих всегда, а более специализированных детёныши чаще всего с пятнистой окраской: на тёмном фоне многочисленные светлые пятна; только у наиболее специализированных оленьих (северный олень, лось) окраска всегда однотонная. У многих оленьих есть "хвостовое зеркало" — светлое пятно на задней стороне ляжек и на крупе. Волосы на нём обычно удлинены, при тревоге топорщатся, увеличивая его площадь, что имеет сигнальное значение.
Череп с относительно слабо пневматизированными костями лицевого отдела. Имеется длинная диастема, т.е. беззубый участок челюсти перед щёчными зубами. Зубная формула I 0/3, C 0-1/1, P 3/3, M 3/3 = 32-34. Верхние клыки обычно редуцированы; у примитивных безрогих оленьих они гипертрофированы (сходно с кабарговыми). Щёчные зубы с низкой коронкой, их жевательная поверхность селенодонтная.
Желудок 4-камерный, разделен на рубец, сетку, книжку, сычуг; жёлчного пузыря у большинства оленьих нет.
В кариотипе 6-74 хромосом; внутривидовой хромосомный полиморфизм редок; у нескольких видов имеется половой тривалент.
Ареал охватывает Евразию, Сев. и Юж. Америку, Сев.-Зап. Африку; некоторые виды интродуцированы в Австралии, Нов. Гвинее, Нов. Зеландии. Обитают главным образом в лесах.
Территориальны; для северных видов характерны сезонные кочёвки. Стадные, на период размножения 
формируют гаремы. Гон сопровождается турнирными боями. Беременность 5-10 мес, в помёте чаще всего 1-2 детёныша (у примитивных видов семейства до 4). Многие многочисленны; некоторые редки, сохранились только в парковом хозяйстве (олень Давида, месопотамская лань); несколько видов истреблено (например, олень Шомбурка).
Все оленьи — объекты промысловной и спортивной охоты: добываются главным образом ради мяса, пантов, трофеев. 1 вид (северный олень) одомашнен. Несколько видов — объекты пантоводства, их разводят в полувольном состоянии специально для срезания пант, из которых делают ценные лекарственные препараты (прежде всего пантокрин).
Читайте также: